Kornilova L.N., Naumov I.A., Glukhikh D.O., Ekimovskiy G.A.,
Sagalovich V.N., Khabarova E.V., Pavlova A.S.
Since ancient times, mankind aware of a feeling of nausea or dizziness caused by the movement – motion sickness (MS). The Aristotle’ diaries describe the unpleasant sensations in soldiers moving on elephants in the army of Alexander the Great. Initially, this condition associated with long swing of the head and torso, but soon empirically humanity came to the conclusion that the cause of MS is ear. With the development of transport, especially shipping and aviation, it became apparent that the reason for the development of the MS is the vestibular apparatus. Severe inhibition or absence of the function of the vestibular apparatus makes people insensitive to the MS. Reinforcement of point of view about the leading role of the vestibular system in the development of the MS is resistance to it individuals with dysfunctional labyrinths in parabolic flights and under the effects of Coriolis’ and precessional accelerations. Deaf people are immune to the MS. It has been established that animals with destructed vestibular apparatus and vestibular pathways in the central nervous system (CNS) are immune to accelerations.
It has been established that the vestibular apparatus in response to acceleration, changes in body position and gravity provides:
- orientation in space;
- balance control;
- image stabilization on the retina;
- mantaining a still image on the retina during head or body movement.
However, implementation of spatial orientation, the accuracy of visual tracking (static and dynamic fixation saccades, gaze holding, smooth pursuit), perfect coordination and balance control needed as proper functioning of individual sensory systems and their coordinated interaction (convergence) in the integrative structures of the CNS. This applies primarily for visual, vestibular and motor systems, which are components of a single perceptive-motor system “body-environment” and are functionally inseparable.
In weightless statoliths lose weight and stop pressure on the membrane, and thus information on changes in head position and linear movement of the body (familiar for terrestrial conditions) does not arrive in the CNS at all or arrives distorted. Thereby, in weightlessness, the information coming from the vestibular system does not coincide with the information coming from other sensory systems, thus violated the usual sensory communication and the brain at the beginning of the spaceflight can not correctly interpret the signals coming into it, which leads to the development of the sensory conflict. The first level of this conflict is the otoliths-canals conflict, then the vestibular-visual, hereinafter – the intersensory disintegration.
Understanding these features of the functioning of the otoliths in weightlessness and the subsequent intersensory disintegration, many researchers foresee the development of sensory disturbances in weightlessness as vestibular dysfunctions. Spaceflight of German Titov and subsequent space missions confirmed the assumptions of researchers.
In the initial period of adaptation to weightlessness and during readaptation to Earth’s gravity almost all cosmonauts noted a number of specific, atypical sensory reactions, in particular orientation illusions, dizziness, difficulty in fixation and visual tracking of objects in field of view. This condition is experienced as uncomfortable, especially in presence of additional vegetative symptoms.
Phenomenological similarity of symptoms developing in weightlessness (microgravity) and during readaptation to Earth’s gravity with terrestrial forms of motion sickness determined the name – SPACE MOTION SICKNESS (SMS).
SMS – is a condition where normal physiological adaptation goes into a phase of decompensation on the level of integrative mechanisms of the CNS, i.e. when adaptive responses of the space adaptation syndrome (SAS) reach the clinical level and are accompanied by deterioration of the professional performance.
Many researchers consider the development of the SMS with intra- and extralabyrinths mechanisms. In our opinion, such division is incorrect, since the vestibular apparatus (because of the vastness of functional connections between vestibular and other sensory systems) obligatorily involved in all integral reactions of the body.
The aims of this study were:
- to determine the nature of the interaction between otolith-oculomotor and cupular-oculomotor subsystems in readaptation to terrestrial conditions after prolonged exposure to weightlessness;
- to study the effects of weightlessness on the characteristics of all forms of visual tracking (smooth pursuit, fixation saccades, gaze holding);
- to define of the role and importance of the vestibular system to provide visual tracking;
- to clarify the relationship between characteristics of the visual tracking and state of the vestibular function;
- to determine the extent of involvement of different levels of locomotion’s innervation mechanisms in the development of visual tracking disorders after spaceflight.
Assessment of the vestibular function and visual tracking were performed using computerized stimulation programs on the hardware-software complexes (HSC) «SENSOMOTOR», «OCULOSTIM», «VIRTUAL» (last one – onboard the ISS during spaceflight), which are based on:
- PC with 2 monitors: one for the presentation of visual stimulation to the cosmonaut, second – for control of physiological responses of the cosmonaut by the surgeon/researcher.
- EOG recorder – a four-channel EOG-amplifier operating on the direct current with a sampling rate of 250 Hz to record eye movements and a three-axis accelerometer combined with an angular velocity sensor with a sampling rate of 100 Hz to record head movements. Accuracy of the recorded and processed EOG data was estimated to be < 0.5°. In our studies eye movements were recorded using both electrooculography (EOG) and videooculography (VOG).
- Joystick for registration of begining, direction, intensity and ending of dizziness and other coordination illusions.
- Software – battery of computerized tests, providing: stimulation (separate, selective, combined and polymodal, strictly dosated and strictly directed) of the visual and vestibular sensory inputs, recording of physiological reactions and responses, centralized storage and processing of the recorded signals.
Processing and analysis of records of eye and head movements was performed using software specially developed by staff of laboratory of Vestibular physiology at the SSC RF – IBMP RAS (Sagalovitch S.V., Stephankov D.V., Azarov K.A., Naumov I.A.).
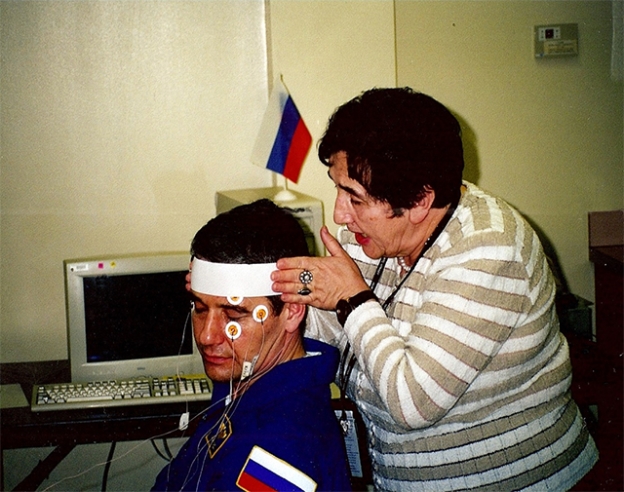
Fig. 1 shows posflight assessment of cosmonauts using the HSC «SENSOMOTOR» and «OCULOSTIM»
 |
 |
Fig. 1. Posflight assessment of cosmonauts using the HSC«SENSOMOTOR» and «OCULOSTIM»
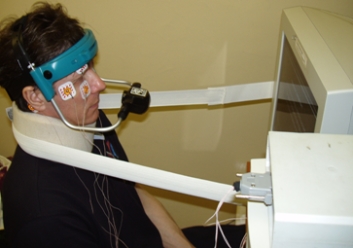
Eye movements were recorded using both electrooculography (EOG) and videooculography (VOG) methods. Eye movements were recorded by VOG-complexes VNG Ulmer (made by Synapsys Europe and Heinemann Medizintechnik GmbH) and Chronos Vision ETD (made by Chronos Vision GmbH) (Fig. 2).

a) |
b) |
Fig. 2. Registration of eye and head movements of cosmonauts: a) ETD, b) VNG Ulmer with EOG and VOG simultaneous recording
To register horizontal, vertical and torsional eye movements subject wear special glasses (helmet) fitted with infrared cameras with scanning frequency of 50 Hz (in the case of VNG Ulmer) and 100-400 Hz (in the case of Chronos Vision ETD). Range of eye movements was recorded up to 55° horizontally and 35° vertically, accuracy of recognition of eye position – 0.25° (VNG Ulmer) and <0.1° (Chronos Vision ETD) [Clarke A. et al.]. Processing of VOG-records and selection of components of eye movements (horizontal, vertical, torsional) were performed by programs developed and built into complexes by manufacturers.
Studies involving the simultaneous use of two methods of recording of eye movements (VOG allows to record torsional eye movementd and EOG allows to record ocular responses with closed eyes) for the first time allowed to assess the state of static and dynamic components of the vestibular function in humans, as well as examine the gaze holding at eccentric eye position vertically and horizontally. Combined use of VOG and EOG gave an opportunity to compare the effectiveness of these two methods in assessments of cosmonauts and in model experiments.
During spaceflights on the orbital stations “Salyut-7″ and “MIR” 31 cosmonaut participated in the study, 27 of whom were in long-term expeditions (76-438 days), and 4 in brief short-term (7-9 days) spaceflights. 13 of 31 cosmonauts have been in spaceflight 2 or more times. Age of cosmonauts was 28-50 years. Each of cosmonaut was examined preflight on L-30 and L-10 days before launch; in spaceflight on days 2(3), 5(6), 28(30) of staying in weightlessness, then once per a month or two months before the end of the flight; postflight on days R+1-2, R+4-5, R+8-9 after landing, and on days R+14 or R+19 in case of need.
Within the space experiment «VIRTUAL» and pre- and postflight science experiment «SENSORY ADAPTATION» there were examined 40 russian cosmonauts – crewmembers of the longterm expeditions on the ISS (starting from the expedition ISS-3). Length of stay on the ISS was between 129 and 215 days with an average duration of exposure to weightlessness ~175 days. Their ages ranged between 35 and 50 yr, with their average age being 41.5 yr.
All cosmonauts had undergone extensive medical examinations (including examination by an ophthalmologist and neurologist), had normal vision with no oculomotor abnormalities, had no known clinical vestibular problems, and were not taking drugs which affect the nervous system. The study protocol (space experiment «VIRTUAL», science experiment «SENSORY ADAPTATION» and clinical-physiological assessment «Vestibular Function Check up») was reviewed and approved in advance by the Bioethics Board of the Institute of Biomedical Problems and Human Research Multilateral Review Board. All cosmonauts gave written, informed consent before participating in the experiment.
Data was acquired twice prior to spaceflight (L-45 and L-30) and after landing on R+1-2, R+4-5, R+8-9, and sometimes R+14-19 where the exact date of examination (for example, R+1 or R+2) depended on landing conditions and health status of the cosmonaut.
The research program of vestibular function includes the following sections:
- Calibration
A standard five-point calibration routine was used, i.e. ±10º left/right, up/down and center fixation sequence. A calculation of the ocular torsion (VOG) was performed by extraction of iris profiles and subsequent cross-correlation against a zero reference frame.
- Spontaneous Eye Movements (SpEM)
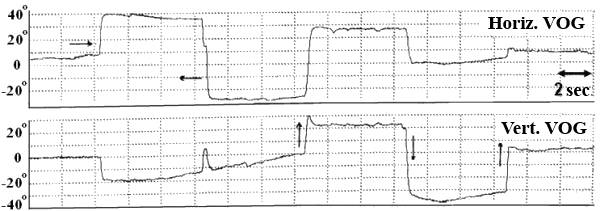
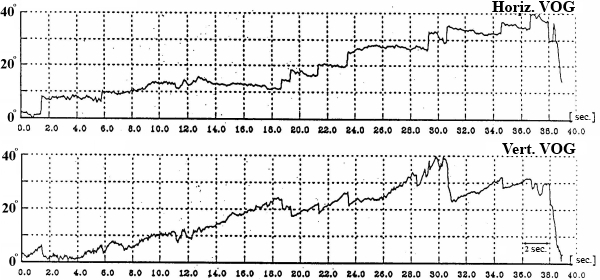
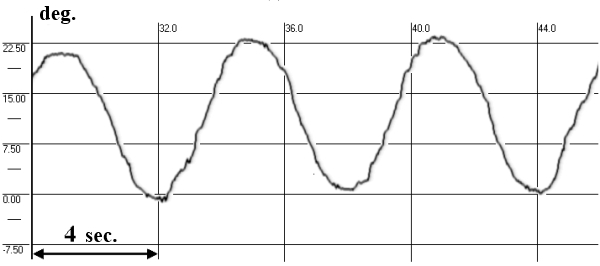
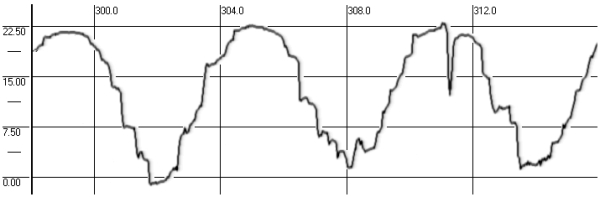
In this test subject held his head strictly vertically (it was fixated by a special head holder) and eyes (closed for EOG and opened for VOG) were positioned initially in the central position of the orbit, and then in response to a sound command were moved to “extreme” right, left, up or down positions and held for 7 seconds in each (Fig. 3). We examined drifts of eyes, periodical spontaneous nystagmus, and gaze evoked nystagmus in end-point positions.
preflight
inflight (3rd day)
inflight (43rd day)
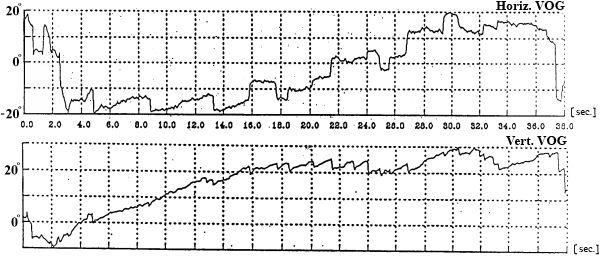
postflight (R+2 day)

Fig. 3. SpEM before, during and after spaceflight
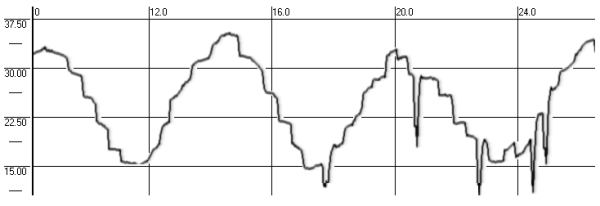
- Static torsional otolith-cervical-ocular reflex (OCOR)
Measurements of the torsional ocular counter-rolling were performed during head tilts to both shoulders (30º tilts controlled by an angle-measuring device – Fig. 4) and head held in these positions for 16 seconds (to prevent dynamical influences on the reflex caused by the head movement). During the test each subject tilted his head twice to the left, and twice to the right shoulder, following a pattern: “center-left-center-right-center…”. OCOR was evaluated by the amplitude of the compensatory torsional ocular counter-rolling.
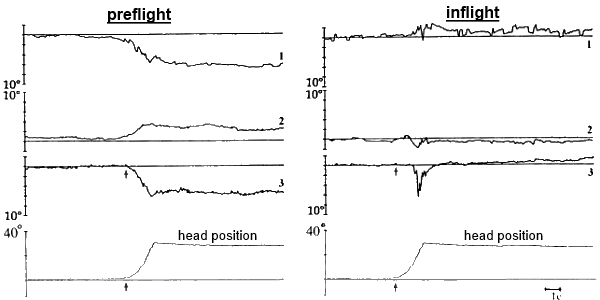
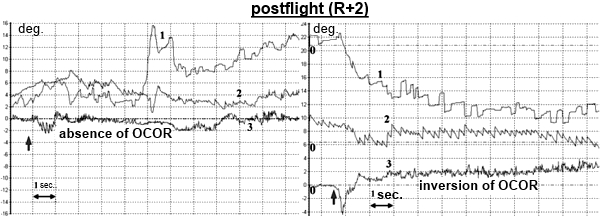
Fig. 4. OCOR before, during and after spaceflight:
1 – horizontal VOG, 2 – vertical VOG, 3 – torsional VOG,
↑ – moment of the head tilt to right shoulder - Dynamic vestibular-cervical-ocular reflex (VCOR)
The subject was asked to make voluntary active head movements in the horizontal (yaw rotation) and sagittal (pitch rotation) plane with a frequency of 0.125 Hz with opened and closed eyes (Fig. 5). To assist the subject a frequency modulated sound signal was used that he could follow. Approximately 7 to 9 shoulder-to-shoulder and up-down cycles were made in each plane.
VCOR was evaluated by amplitude and gain of the compensatory eye movement (gVCOR) – a ratio between velocity of head movement and velocity of the compensatory eye movement.
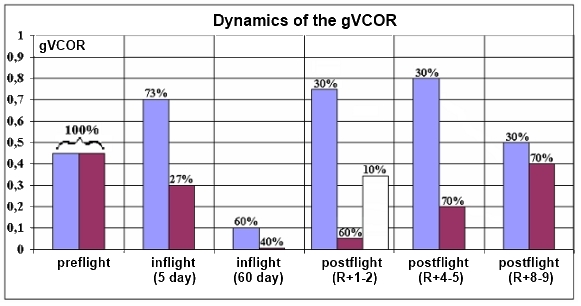
Fig. 5. Dynamics of the gVCOR before, during and after spaceflight
- Vestibular reactivity (VR)
VR and VCOR were examined during the same rotation tests. The difference lies in parameters being evaluated. VR was evaluated by characteristics of the nystagmus which accompanied the mentioned compensatory eye movement (Fig. 6 and 7). The presence of single nystagmus stikes testified normal VR. Strengthening of nystagmus indicates increased VR.
Fig. 6. Examination of the VCOR and VR
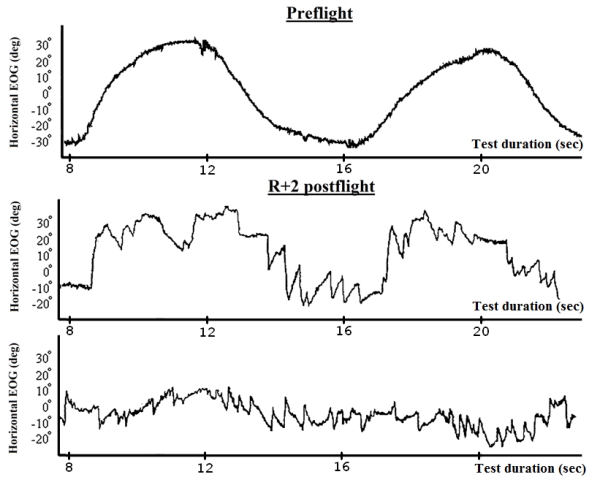
Fig. 7. Vestibular-cervical-ocular reactions in different cosmonauts before and after spaceflight
Visual tracking:
In studying of visual tracking cosmonaut’s head was fixed motionless by a custom-fitted head holder; the distance between the cosmonaut and display was 50 cm. Special optical filters were used to avoid both parasitic glow from the screen and plume from the moving target. The foveal stimulus (visual target) consisted of a small white square (angular displacement less than 1°) that was displayed on the monitor both with and without additional retinal optokinetic stimulation (ROKS) – blurry spots (ellipses), different in shape and diameter, displayed on the screen background and moving in a certain law of motion at ~ 6-9 deg / sec (Fig. 8).
All tests were carried out in a completely darkened room; cosmonauts were given ; 2-3 min for initial adaptation to darkness. Gaze holding test was performed after at least 10-12 min from the beginning of the examination.
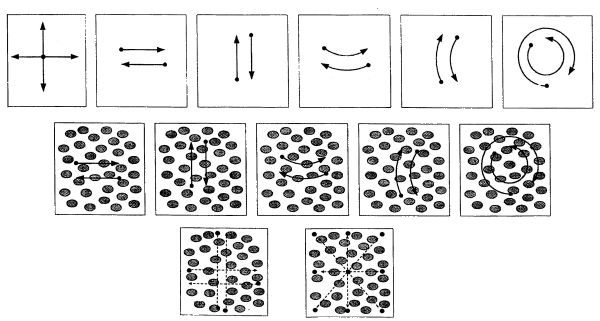
Fig. 8. Visual stimuli used in the study
- Fixation static saccades
This test required subjects to acquire the visual stimulus which was moving saccadically by 20° and held static for 2 s in both horizontal and vertical planes. Approximately 12 to 16 trials (iterations) in each plane were made (Fig. 9).
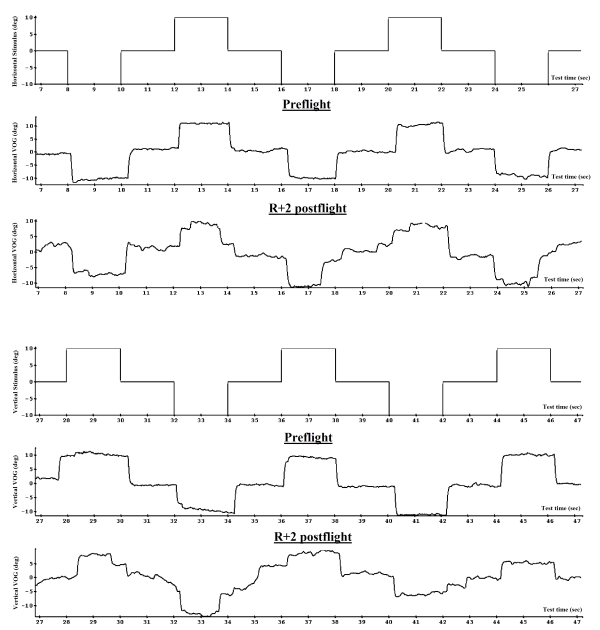
Fig. 9. Fixation static saccades before and after spaceflight (VOG)
- Dynamic saccades
Foveal stimulus initially appeared at the edge of the screen and then was moving to the opposite edge with a constant velocity of 5.7 °/s with an abrupt return in itial position (foveal optokinetic nystagmus). Total displacement of the stimulus was 20°. Approximately 12 to 16 trials were obtained in each plane (Fig. 10).
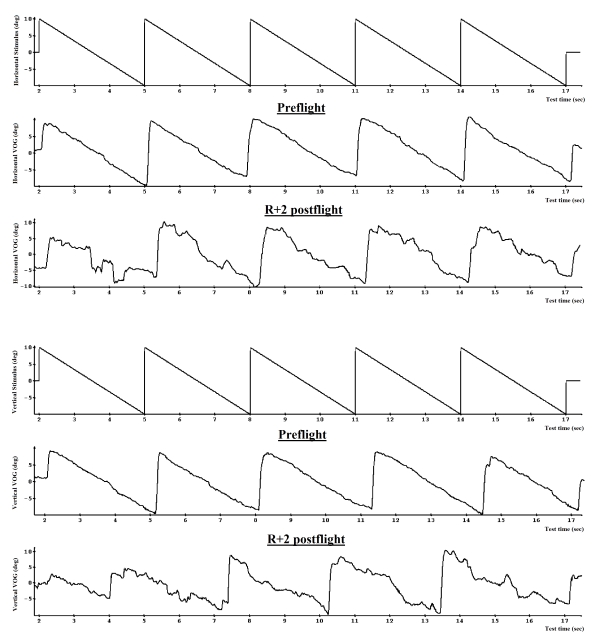
Fig. 10. Dynamic saccades and linear smooth pursuit before and after spaceflight (VOG)
- Smooth pursuit (SP)
Linear SP was examined in the same test with dynamic saccades, sinusoidal SP required the subject to fixate and pursue the target, which moved smoothly and pendularly at a frequency of 0.33 Hz in both horizontal and vertical planes at the same displacement of 20° (Fig. 10 and 11).
preflight
postflight (R+2 day)
postflight (R+5 day)
postflight (R+9 day)
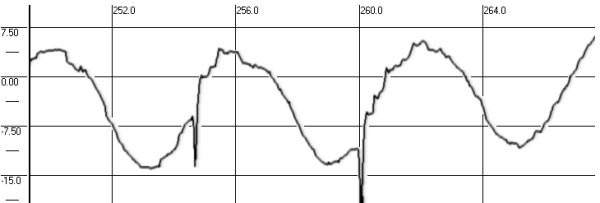
Fig. 11. Smooth pursuit before and after spaceflight (VOG)
- Gaze-holding (GH) on a real and imaginary target
In the beginning of the test the stimulus was randomly displaced by 10° in either the horizontal or vertical plane and illuminated on the monitor for approximately 1 s. The subject was required to acquire the target with a saccadic eye movement. After 1 s the target was blanked. The subject maintained gaze on where the target was before blanking. Then after 9 s a sound command instructed the subject to return his gaze to the central position and wait until the next randomly directed target appeared (Fig. 12).
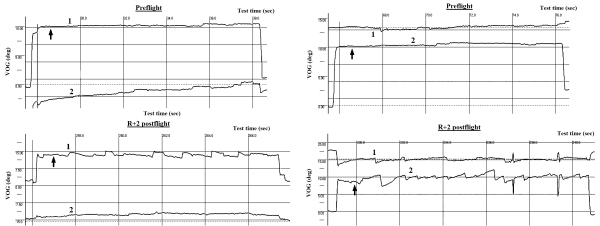
Fig. 12. Gaze-holding on a real and imaginary target before and after spaceflight:
1 – horizontal VOG, 2 – vertical VOG
↑ indicates the moment when stimulus disappeared
For each parameter characterizing the state of vestibular function and visual tracking there were evaluated various statistical characteristics (mean, variance, coefficient of variation etc.). Quantitative assessment and comparative analysis of the parameters were carried out by parametric and nonparametric methods of variance and correlation analysis. In all cases when statistical hypnotizes were used (normality of the distributions, homogeneity (equality) of variances, the reliability (significance) of differences) critical significance level α was 0.05.
Results shown in this study demonstrate that most parameters, which characterize the state of the vestibular and oculomotor function during readaptation to terrestrial gravity, undergo significant changes in comparison to the baseline. As we are continuing to observe, unique methods or strategies for gravitational compensation are adopted by individual astronauts and cosmonauts. For the most part we believe that earlier data, as well as data presented here, show that these strategies are not conscious decisions; rather they represent the way that an individual’s CNS adapts to changes that affect performance and function. Along with unique adaptive changes to the novel gravitational environment, we were able to fi nd regular, typical consistent changes that appeared across individuals.
The postflight increase in SpEM, degradation of the GH ability, and the new saccadic tracking strategy instead of the SP indicate the involvement of central mechanisms of the vestibular system and reflect, probably, changes in functioning of the vestibular nuclei, reticular formation of the middle brain, and the cerebellum.
Torsional ocular counter-rolling induced by a body or head tilt was examined after spaceflight as an indicator of the adaptation of otoliths to microgravity. In microgravity there is no otolith-induced compensatory torsional ocular counter-rolling in response to a sustained tilt. Previously it was shown that torsional ocular counter-rolling was reduced both after short-duration and long-duration missions. Similar changes were observed in two primates after spaceflight [Cohen B.]. However, there was no change in torsional ocular counter-rolling in astronauts who flew in the 1998 Neurolab Mission (STS-90) [Clement G., Moore S.]. In this experiment four crewmembers were examined pre-, in-, and postflight. Astronauts were exposed to artificial gravity (1 g or 0.5 g centripetal acceleration) generated by centrifugation. Absence of differences between pre-, in-, and postflight torsional ocular counter-rolling could be explained by an intermittent exposure to artificial gravity during the 16-d mission, which had prevented deconditioning of otolith-ocular reflexes in microgravity. These data allow us to consider in-flight centrifugation as a countermeasure for vestibular disorders occurring during and after spaceflight.
Our postflight data (absence, inversion, or a sharp decrease of the OCOR) show that adaptation to microgravity is associated with a deep and long suppression of OCOR.
Postflight changes in OCOR are consistent with results of histological studies in rats flown in space [Krasnov I.B.]. There is morphological evidence for hypofunction of receptor cells in the utricle, a decreased afferent input to the vestibular nuclei, and a decreased vestibular input to the cerebellum flocculus. Morphological studies of multipolar neurons in the reticular formation demonstrated disturbance of the evolutionally formed intersensory relations within the CNS, and establishment of new relations to allow spatial orientation in microgravity. Rearrangement of intersensory relations (neuron axons of the vestibular nuclei complex) after spaceflight takes some time, which explains the absence of the OCOR in several cosmonauts tested shortly after landing. Reconstruction of this reflex on return to the Earth takes a certain time during which there is readaptation of otoliths to terrestrial conditions.
Statistical analysis showed that for both leftward and rightward head tilts OCOR on R+1-2 and R+4-5 was significantly decreased in comparison with the baseline and R+8-9, and also there was a significant difference between R+1-2 and R+4-5. OCOR reached its preflight level only on R+8-9.
Changes in gain of horizontal vestibular-ocular responses (head yaw rotations) point to discoordination of eye and head movements. gHVCOR was increased in some cosmonauts (35%) and decreased down to zero in others (46%); 19% of cosmonauts had gHVCOR equal to the baseline. The decrease in horizontal gHVCOR is largely an outcome of suppression of the compensatory torsional ocular counter-rolling by a still existing central deafferentation of the otolith signal in some cosmonauts during the first days postflight. In the work [Cohen B., Dai M.] dedicated to bio-satellites it was shown that during spaceflight primates have horizontal gHVCOR reduced to 15 – 50% and this suppression occurred until R+11.
The sharp rise in intensity of the vestibular nystagmus during head yaw rotations was observed in many cosmonauts after landing, implying an increased dynamic excitability of the vestibular input. Earlier investigations of vestibular receptors of frogs [Gualtierotti T.] had also shown increases in activity of the dynamic component. Some static cells of vestibular receptors were also found to function dynamically in microgravity
Observed sensorimotor disorders in microgravity are usually accounted for by the development of the sensory conflict, the first level of which is the otoliths-canal conflict. In terrestrial conditions, in most cases the structures of the receptors are in reciprocal relations but some individuals have synergetic relationships between otoliths and semicircular canals.
In microgravity there is no natural stimulation for otoliths and thereby their functioning decreases significantly. We believe that in cases of reciprocal relationship, semicircular canals on the contrary will function with increased excitability. However, in the case of the synergic nature of the otoliths-canal relationship, the canals’ excitability will be decreased.
On return to Earth, taking into account reciprocity of the otoliths-canal interaction, reactions of the semicircular canals should have been dampened. Yet, the picture is opposite. This can be ascribed to either a synergic interaction of the otoliths and semicircular canals after flight, or, which seems more probable, persisting central deafferentation of the otolith input and reciprocal otoliths-canal interaction at the beginning of readaptation.
Comparative analysis showed increase of the dynamic and decrease of the static vestibular excitability in the majority of cosmonauts during the first days postflight. The revealed negative correlation between OCOR and VR could point to a reciprocal nature of the otoliths-canal interaction in the studied group of cosmonauts. Similar relationships between otoliths and semicircular canals were noted in model experiments on Earth. However, microgravity can affect otolith function and semicircular canals not only directly (because of the lack of gravity) but also mediately because of the support unloading and, thus, minimization of the support and tactile-proprioceptive afferentation. It occurs via the central integration multisensory structures of the CNS which are sites of the convergence of afferent signals of varying sensory modalities: visual, vestibular, tactileproprioceptive, support. Thus, in microgravity we should take into account both changing of sensitivity of the vestibular system and removing of inhibitory and modulatory effects of the support and tactile-proprioceptive afferentation. In research studies carried out in a model-based microgravity (“dry” immersion) appearance of the spontaneous nystagmus was found in the first day after the experiment. After immersion we found changes concerning a peripheral part of the vestibular system: absence or inversion of the OCOR and positional nystagmus with an inverted reflex.
It is generally believed that visual input is a source of reliable information on Earth or in altered gravity, as it does not belong to a gravity-dependent system, but is rather an organ for perception of visual objects. Indeed, previous results of cosmonaut ’ s postflight medical examinations indicate preservation and clinical wellbeing of the main visual functions. However, since on Earth the vestibular system plays the leading role in realization of oculomotor responses insuring accuracy of the pursuit of visual objects, in weightlessness the above changes of the vestibular function will influence visual tracking as well.
Comparative studies of significance and the role of the vestibular and support afferentation in generating gaze control in microgravity have shown the leading part of the vestibular system in visual tracking. Observable changes in visual tracking were first of all caused by the vestibular deprivation which was developing in microgravity (functional deafferentation of the otoliths sensory input) since it was shown that additional vestibular stimuli (active head rotations) significantly improved the characteristics and parameters of the visual tracking. Exactly by additional stimulation/centrifugation of the vestibular input we could explain the lack of changes in ocular counter-rolling in microgravity in the work of [Moore et al.]
After spaceflight the precision (amplitude), velocity and time parameters of visual tracking were degraded almost in all cosmonauts. During the whole readaptation period there was a statistically significant decrease in amplitudes and peak velocities of both horizontal and vertical saccades, as well as decrease in gain of horizontal and vertical smooth pursuit.
The largest changes in the visual tracking (persistent corrective saccades during gaze fixation, breakdown of the SP reflex and its replacement by a saccadic tracking) were observed in cosmonauts who, along with exaggerated reactivity of the peripheral vestibular input, also developed central changes in the oculomotor system (i.e., square wave jerks, slow wave drifts in SpEM, gaze-evoked nystagmus, spontaneous nystagmus). Deep disturbances in the innervation mechanisms of eye movements were demonstrated by development of the new tracking strategy: cosmonauts approached and tracked stimulus using a sequential set of saccades (even during a smooth pursuit task). This new strategy resulted in significant increase of the total time (up to 2-3 times) required for initial consideration and recognition of the target and further gaze holding. This replacement of the smooth pursuit by a saccadic one can be stipulated by a greater stability of the saccades innervation mechanisms. Saccadic eye movements are implemented by evolutionary older structures of the brain and have a reflexive nature whereas SP is executed by younger parts of the cerebral cortex.
It should be noted that in most cases horizontal eye movements were more stable and precise than vertical. From the standpoint of human evolution this appears reasonable since visual objects that surround us usually move horizontally. Furthermore, we seldom move (other than locomotion) in the vertical plane. Analysis of available neurophysiologic and clinical literature reinforced the statement that instability of vertical eye motion as compared to the horizontal one might be due to the difference in location of the nerve pathways controlling these motions. Horizontal gaze stability is facilitated by more numerous morphological elements leading from the oculomotor retinal neurons to cortical areas.
Although vertical eye movements are more sensitive to altered gravity, as opposed to the horizontal visual tracking the vertical one was not correlated with the state of the vestibular function. This once again indicates that horizontal and vertical eye movements are realized by different neural mechanisms.
Changes in the nature and pattern of correlations between studied parameters and increases in their variability reflect instability in postflight sensorimotor adaption to normal terrestrial gravity
It was shown that disturbances and disorders in visual tracking were observed in cosmonauts who along with decreased tonic (static) and increased dynamic vestibular excitability also had various central changes in the vestibular system and some changes in the nature of the relationship between the vestibular system and other subsystems of the CNS.
According to results obtained, at the beginning of readaptation to the Earth ’ s gravity, the majority of cosmonauts have shown:
- increase of vestibular reactivity;
- suppression of the otoliths function and low tonic (static) vestibular excitability;
- reorganization of the interaction between otoliths and semicircular canals; and
- changes in central mechanisms of the intersensory interactions resulting in a significant decrease of amplitude and velocity parameters of visual tracking, and development of the saccadic tracking strategy instead of the smooth pursuit.
As a rule, during the first week of readaptation a sharp decrease in tonic (static) vestibular excitability (OCOR) was accompanied by an increase of the dynamic one (VR). Furthermore, the negative OCOR/VR correlation suggests that there is a reciprocal nature of the interrelation between these reflexes in cosmonauts.
Microgravity has no direct effect on gaze control; however, by remodeling the vestibular afferentation and central mechanisms it did affect all forms of visual tracking. The established disturbances in precision of the saccadic function and smooth pursuit (particularly in the vertical plane) and adoption of the new “saccadic” tracking strategy prolong substantially (up to 2-3 times) the total time required to acquire, recognize, and to fixate gaze on the target. These structural changes in the visual tracking function were typically seen in cosmonauts who in addition to the changes in the vestibular function also developed spontaneous nystagmus, gaze nystagmus, slow deviation eye movements, and square wave jerks. It indicates that these disorders were driven by alteration of the central mechanisms of intersensory interactions rather than by a peripheral part of the vestibular system.
At least until R+8-9, measured parameters tended toward, but did not fully regain their baseline values. These observable changes in the vestibular and oculomotor systems are a typical adaptive response of sensory systems to an altered gravity, and require time to rearrange intersensory relationships for the Earth ’ s gravity. It also appears that the adaptive response may be related to the amount of time spent in spaceflight.
Not one test but a whole battery of computerized tests (which provides both separate, election and combined, strictly dosated and directed stimulation of various sensory inputs) has revealed variety of different types of disturbances in the eye movements controlled both by the vestibular system, and by a complex hierarchy of innervation mechanisms disposed at all levels of the central nervous system. Postflight diversity of disorders indicates that in thier formation there are involved all levels of the vestibular and oculomotor innervation mechanisms.
Our results indicate the need to develop countermeasures for interplanetary spacefl ight. If no appropriate countermeasures are taken during future Moon or Mars missions, cosmonauts may encounter problems on detailed tasks involving visual tracking and fixating, regardless of the disorientation that they are sure to encounter.
The authors are thankful to the cosmonauts for their participation in examinations, and to flight surgeon, Alexander V. Vasin, M.D. (Yu. A. Gagarin Cosmonaut Training Centre, Star City) for his help during the experiments.