THE EFFECTS OF THE DRY IMMERSION ON VESTIBULAR-OCULAR REFLEXES AND VISUAL-MANUAL TRACKING
Kornilova L.N., Glukhikh D.O., Naumov I.A., Khabarova E.V.,
Ekimovskiy G.A., Sagalovich V.N., Pavlova A.S.
Organizer of the immersion experimental base at the SSC RF – IBMP RAS and the leader of all immersion experiments is the head of Department of Sensorimotor Physiology and Prophylaxis Prof., Dr.Med.Sci., corresponding member of the Russian academy of sciences Kozlovskaya I.B.
Quite a number of works have been devoted to the study of coordination and synchronization of hand and eyes movements during pursue of visual targets in terrestrial conditions.
On Earth the process of perception and spatial orientation, visual (VT) and visual-manual tracking (VMT) are determined by multimodal activities of the gravity-dependent and independent afferent systems and by the nature of their intersensory integration. Established multimodality of the afferent provision of perception and spatial coordination is the basis of reliability and adaptive capacity of the VT and VMT.
In altered gravity (weightlessness) vestibular system (otoliths receptors) is the trigger of abnormal vestibular reactions and sensorimotor disorders, which may lead to reduced accuracy of the VT and VMT. Disorders in kinematics of hand movements in weightlessness were found previously in some spaceflight experiments.
However weightlessness can affect the otolith function and function of the semicircular canals both as a direct result of the loss of the gravitational effects on otoliths as well as indirectly through a multi-sensory integrative structures of the central nervous system (CNS), where the convergence of afferent signals of different sensory modalities takes place, by eliminating the support and minimizing the tactile and proprioceptive afferentation.
To understand the role and the specific contribution of vestibular and support inputs in formation of intersensory interaction, in ensuring the accuracy of the VT and VMT there are necessary ground simulation (model) experiments (Fig. 1).
 |
 |
 |
 |
Fig. 1. Horizontal “dry” immersion experiment
Previously it was shown that of all experimental simulation models used on Earth, the most adequate from the point of view of the analogy of oculomotor reactions to reactions observed in weightlessness, is immersion [Kozlovskaya I.B., Kornilova L.N. et al.]. Immersion itself does not directly affect the visual and vestibular receptors, but it creates the support unloading and minimizes muscle activity, i.e. in other words, removes the support and reduces the proprioceptive afferentation in integrative structures of the CNS. In turn, these structures is the place of the convergence of multi-sensory afferent signals of different sensory modalities, primarily visual, vestibular, motor and support. It can be expected that sensory deprivation caused by the removal of support and minimization of muscle activity in immersion leads to a change in the functioning of the vestibular nuclei and multi-sensor activating structures of the midbrain.
Changed functioning of the multimodal vestibular nuclear complex caused by removal of the support afferentation can affect characteristics of the VT and VMT because of the inseparable phylogenetically formed connections between the vestibular, oculomotor and motor nuclei.
The purpose of the study was to examine the role of the support-proprioceptive factor in the functioning of the vestibular system, in the mechanisms of intersensory interaction and, in particular, visually-mediated interaction between hands and eyes motor systems.
The objectives of the study were:
- to study the effects of decreased support-proprioceptive afferentation in horizontal “dry” immersion on characteristics of the VT and VMT;
- to evaluate the accuracy and synchronization of the “eyes-hand-stimulus” motor coordination in immersion while pursuing moving visual stimuli without additional (optokinetic) visual noise;
- to study the effects of decreased support-proprioceptive afferentation on nature of vestibular-ocular reflexes;
- to clarify the relationship between accuracy of the VT/VMT and state of the vestibular function (VF).
The study of the effects of immersion on characteristics of VMT involved 30 male subjects, aged from 19 to 31, which passed a detailed medical examination, were found to be healthy and admitted to the experiment. 16 subjects participated in the 7-day immersion, 14 subjects – in the 5-day immersion.
Examinations of the VMT were carried out twice before immersion (L-7 & L-3) (Fig. 2), in 3-4 hours after diving in the immersion bath (I-1), on 3 and 5-6 days of staying in the bath (I-3 & I-5-6), in first hours after pulling out from bath (R+1), on 3-4 and 5-6 days after immersion (R+3-4 & R+5-6).

Fig. 2. Examination of the VMT before immersion (baseline)
During the study of VMT eye movements were recorded by electrooculography (EOG) approach using the hardware-software complex “OCULOSTIM – CM”.
Signals of hand’ motor act were recorded in two axes using the Saitek Cyborg Evo joystick. The specificity of the tests used in examinations and the presence of visual bio-feedback for hand movements (there was a visible target on the screen which represent the current angle (position) of the joystick) allowed us to avoid specific calibration procedures (the range of values of the joystick’ axes was scaled to the area of ±15º and thus both axes of the joystick registered hand movements in degrees).
ПPosition of the joystick during the whole experiment was identical: joystick’ handle was located opposite to the dominant hand of the subject, providing the 90º angle of the elbow. These experimental conditions were abided by all subjects, thus ensuring identical conditions for all participants.
Examination contained computerized stimulation programs (tests), which were presented to the subject using a virtual reality glasses – i-glasses PC 3D (i-O Display Systems, Sacramento, USA)
Examinations of the VT and VMT before and after immersion were carried out in horizontal (lying) position on the bed (Fig. 2).
Examinations of the VMT contained following tests, which were carried out in a darkened room after 7-10 minutes adaptation to horizontal position:
- EOG calibration (in the start and end of each examination). Fixation eye movements during pursue of the jerky (saccadically) movement of the foveal stimulus (white target with a size of 1º) from central position to horizontal and vertical sides in a range of ±10º;
- visual-manual tracking (VMT) with a visual bio-feedback for hand movements. Two visual targets were demonstrated on the screen: one (“test” stimulus) was moving independently of the hand (joystick) movements according only to the stimulation program of the test, another target (“bio-feedback” stimulus) was moving by the subjects’ joystick. Subject was asked to combine (“catch”) both targets when “test” stimulus was moving a) saccadically in ±10º range on horizontal and vertical planes, b) smoothly (linearly and sinusoidal on horizontal and vertical planes) with a 0.16 Hz frequency in ±10º range, and circularly counter- and clockwise with a 0.08 Hz frequency in 20º range.
To analyze the VMT there were evaluated time, amplitude and velocity characteristics of the eyes and hand movements, and their accordance with the stimulus movement. Including efficiency ratios (eVT and eMT) and gains of visual and manual tracking (gVT and gMT) as ratios of respectively amplitudes and velocities of eyes/hand movements to the stimulus movement, percent (%) of corrective saccades and corrective hand movements.
Processing and analysis of oculomotor and hand signals were made using specifically developed software programs and scripts (Naumov I.A., Khabarova E.V., Azarov K.A.) with mathematical software package Mathworks Matlab.
The criterion of adequacy of VT and MT was that their time and amplitude-speed performance characteristics corresponded to the characteristics of the visual stimulus.
Vestibular function (VF) was evaluated by accuracy of perception of subjective visual vertical (SVV) as well as by nature of vestibular-ocular reflexes recorded using electro- and videooculography (VOG) approaches before (L-7, L-3) and after immersion (R+1, R+3-4, R+5-6).
Perception of the SVV was evaluated both in sitting position and lying on left side (left lateral decubitus) (Fig. 3). In the SVV’ test we evaluate the angle (error) between the line on the screen which was freely controlled by the subject with a mouse controller and true (gravitational) vertical. Test was carried out in complete darkness without any orienteers.
 |
 |
Fig. 3. Examination of perception of the SVV lying on left side
VOG was used to examine following vestibular-ocular reactions:
- Spontaneous eye movements (SpEM), which were examined with a straight vertical position of the head, fixated motionlessly. There were evaluated floating (drift) and saccadically eye movements, spontaneous nystagmus, gaze-evoked nystagmus in central position and in right, left, up and down positions when eyes were moved by sound command. Gaze in each position was held for 7 sec.
- Static torsional otolith-cervical-ocular reflex (OCOR)which was determined by amplitude of the compensatory torsional counter-rolling after 30° head tilts to the left and right shoulders. To avoid the influence of dynamic effects on the OCOR after each movement (tilt) of the head it was held motionlessly for 16 sec. The angle of the tilt was controlled by specifically-developed inclinometer (Fig. 4).

Fig. 4. Examination of the OCOR
In examinations we used head tilts with and without gaze fixation on a distant visual target. Both approaches showed almost identically results in terms of amplitude of compensatory torsional counter-rolling (in some cases the difference was in range of ±0.5° because of the systematical error in procedure of cross-correlation recognition of the iris segment), but test with gaze holding contains additional valuable information – nystagmus, which could be depressed by fixation reflex. Thereby, we analyzed OCOR test with head tilts without gaze fixation.
- Dynamic vestibular-cervical-ocular reactions (VCOR)), which were evaluated by gain (gVCOR) as ratio between velocity of the compensatory eyes counter-rolling and velocity of head rotations around the longitudinal axis of the body with a frequency of 0.125 Hz without gaze fixation.
- Vestibular reactivity (VR), which was evaluated by duration and intensity of the vestibular-invoked nystagmus, which was overlapping (stratifying) the dynamic horizontal counter-rolling of the eyes (during the VCOR test).
To register horizontal, vertical and torsional eye movements we used VOG-complex Chronos Vision ETD (Berlin, Germany) (Fig. 5). Subject wore a special helmet, equipped with infrared video cameras. The range of registered eye movements is up to 55° horizontally and 35° vertically. A sampling frequency was 200 Hz (200 frames per sec.). VOG records was processed using Iris Tracker software (part of ETD-complex) with a measurement resolution < 0.05º in all three (horizontal, vertical and torsional) components.
 |
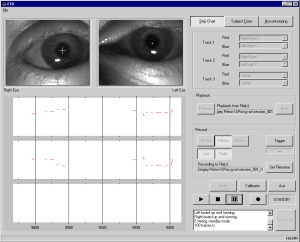 |
Fig. 5. VOG-complex Chronos Vision ETD
A standard five-point calibration routine was used, i.e. 10º left/right, up/down and center fixation sequence. A calculation of the ocular torsion was performed by extraction of iris profiles and subsequent cross-correlation against a zero reference frame.
To analyze VOG there were evaluated: amplitude of the torsional compensatory counter-rolling in a static (held) head position after the tilt (amplitude was evaluated on 10-14 sec after the tilt), gain of OCOR (ratio between amplitude of the torsional counter-rolling and angle of the head tilt – gOCOR), gain of VCOR (ratio between velocity of the compensatory eyes counter-rolling and velocity of head rotations – gVCOR), spontaneous and vestibular-invoked nystagmus (time, amplitude and velocity characteristics).
For each parameter characterizing the state of VMT and VF there were evaluated various statistical characteristics (mean, variance, coefficient of variation etc.). Quantitative assessment and comparative analysis of the parameters were carried out by parametric and nonparametric methods of variance and correlation analysis. In all cases when statistical hypnotizes were used (normality of the distributions, homogeneity (equality) of variances, the reliability (significance) of differences) critical significance level α was 0.05.
To check the significance of differences between values before, during and after immersion following methods were used:
- F-test (ANOVA) with multiple pairwise comparisons made by Tukey’, Dunnet’ and Newman-Keuls’ tets;
- Friedman’s criterion with multiple pairwise comparisons made by Wilcoxon test with Bonferroni correction.
Normality of distributions was checked with Lilliefors’ tests, homogeneity of variances – with Levene’ tests. The presence or absence of pair correlations between the studied parameters were tested using Pearson’s and Spearman’s correlation coefficients.
The whole statistical analysis was made using mathematical software packages Mathworks Matlab и SPSS Statistics.
The study of effects of support-proprioceptive deprivation on characteristics of the visual-manual tracking (VMT)
Accuracy of the VMT:
Accuracy of the VMT was evaluated by efficiency (amplitude) ratios of the visual and manual tracking (eVT and eMT respectively) before, during and after immersion. Dynamics of the eVT and eMT is presented in Fig. 6.

Fig. 6. Dynamics of efficiency ratios of visual (eVT) and manual tracking (eMT):
* – significant difference from the baseline for VT (p < 0,05),
# – significant difference from the baseline for MT (p < 0,05)
As it can be seen from the figure, significant changes of eMT (in comparison with baseline – values before immersion) were found only on I-1 and I-3 days during immersion. Analysis of eVT has shown that significant decrease of the eVT was observed throughout the whole immersion and on R+1 and R+3 days after immersion. eVT returned to the baseline only on R+5-6 day after immersion.
Group and individual coefficients of variation (CV) of amplitudes of MT were more than 3 times lower (3-6% during the whole experiment) than CV of amplitudes of VT (~2% before and 10-20% during and after immersion). These data indicate a slight variation and high stability in performance of MT, which was significantly higher than accuracy of VT.
Velocity of the VMT:
Velocities of the VMT were evaluated by gains of visual and manual tracking (gVT and gMT respectively) as ratios of velocities of eyes/hand movements to velocity of the stimulus movement.
Fig. 7 shows that velocity of MT before, during and after immersion was close to velocity of the stimulus. Statistical analysis found no significant differences in gMT in comparison with baseline and values after immersion.
Analysis of VT found significant decrease of the gVT in comparison with baseline on I-3 and I-5-6 days during immersion and on R+1 and R+3-4 days after immersion. It should be noted that on I-1 day during immersion gVT was 10-15% lower than the baseline, but because of the high variability of samples, a statistically significant difference between them was not found.

Fig. 7. Dynamics of gains of visual (gVT) and manual tracking (gMT):
* – significant difference from the baseline (p < 0,05),
As in the case of analysis of amplitudes, group and individual CV of velocities of MT were much lower than CV of VT velocities (~4-8% for MT and ~15-24% for VT). It indicates that characteristics of the MT in contrast to the performance of the VT more closely correlated with the movement of the stimulus (hand works more precisely than the eyes).
Native curves, which characterize visual and manual tracking of the sinusoidal and linear stimuli, are shown in Fig. 8.

Fig. 8. Fragments of visual-manual tracking’ native curves before and on first day during immersion:
А – sinusoidal movement of the stimulus,
B – linear movement of the stimulus,
1 – eye movement (EOG), 2 – hand movement, 3 – stimulus
Fig. 8 clearly shows a more adequate manual tracking of a moving stimulus in comparison with visual tracking when eyes make a large number of corrective movements. A similar pattern of native curves, observed in immersion, was recorded during first days after immersion in tracking of sinusoidal and linear stimuli.
Latency of the VMT
Fig. 9 shows dynamics of changes in latency of the visual and manual tracking during the whole experiment.

Fig. 9. Dynamics of changes in latency of visual-manual tracking
Analysis of the latency of oculomotor and manual reactions during smooth pursuit of a visual stimulus has shown that it was almost equal to the baseline and did not go beyond the physiological norm: ~0.65 sec – for manual tracking and ~0.25 sec – for visual tracking.
Moreover, subjects often made outrunning movements both with hand and eyes due to the predictable and cyclical nature of stimulus motion.
Latency of the MT in task of fixation and holding a “hand” target on the jerky (saccadically) moving stimulus was more than 1.5-2 times higher than latency of the VT in the same task.
Total reaction time of the VMT
Analysis of the total reaction time of VT and MT has shown that total reaction time of MT was significantly higher in comparison with baseline only on I-3 day during immersion. As concerns VT, there is a replacement of smooth pursuit for saccadic, and total reaction time of VT was significantly higher in comparison with baseline during the whole staying in the immersion bath and in first days after immersion.
The study of effects of support-proprioceptive deprivation on characteristics of the vestibular function (VF)
Spontaneous eye movements (SpEM)
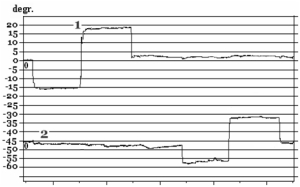
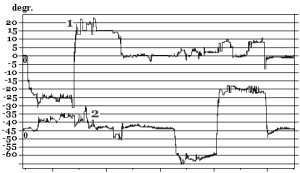
Before immersion all subjects have stable eye movements: there were no drifts and jerky (corrective saccades) eye movements both in the central and sides positions of the gaze. Native curves (VOG) of SpEM before and after immersion are shown in Fig. 10.
before immersion
after immersion (R+1 day)
 |
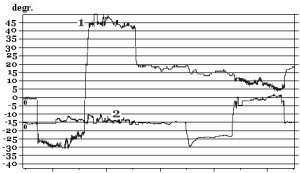 |
Fig. 10. SpEM before and on R+1 day after immersion:
1 – horizontal VOG, 2 – vertical VOG
On R+1 day after immersion two subjects who participated in a 7-day immersion and one subjects from a 5-day immersion experiment had spontaneous nystagmus and gaze-evoked nystagmus (amplitude of fast phase of nystagmus АNy = 4.4±0.7°; velocity of slow phase of nystagmus VNy = 4.9°/s; frequency of nystagmus FNy = 0.5Hz; relative duration of nystagmus in % as a ratio between duration of nystagmus and duration of the whole test TNy = 14.3±4.8%).
On R+3 day after immersion SpEM matched baseline.
Static torsional otolith-cervical-ocular reflex (OCOR)
Before immersion OCOR was adequate on all subjects. Amplitudes of compensatory torsional counter-rolling were symmetrical (for leftward and rightward tilts) and within physiological norm (4-8º). However, one subject showed amplitudes of OCOR decreased to 2.6º for the rightward and decreased to 2.4º for the leftward head tilts, and three subjects showed an asymmetrical reflex (the asymmetry was in the range of 24.8±5.6%).
After immersion all subjects were divided in 4 groups in accordance with their OCOR. In 1-st group (14 subjects) OCOR was within physiological norm and symmetrical during the whole experiment. In 2-nd group (10 subjects) OCOR was significantly decreased (from 6-8º to 2º) on R+1 day after immersion. In 3-rd group (2 subjects) OCOR was increased on 2º in comparison with baseline but still was within physiological norm. In 4-th group (4 subjects) there were atypical forms of OCOR after immersion.
For two subjects from last group on R+1 and R+3 days after immersion head tilt to the shoulder initially caused compensatory ocular counter-rolling in direction opposite to the tilt’ but in a few seconds eyes returned to the zero position and was there during the whole trial (absent of otolith reflex). For another two subjects eyes after initial compensatory ocular counter-rolling rolled in the same direction as tilt (opposite to the initial counter-rolling – inverted reflex). Also all four subjects from this last group had positional nystagmus when head was in the static (motionless) position.
On R+3 day after immersion for all subjects except two from 4-th group OCOR returned to the baseline.
Fragments of the OCOR native curves before and after immersion on some subjects from 4-th group are shown in Fig. 11.
before immersion

after immersion (R+1 day)
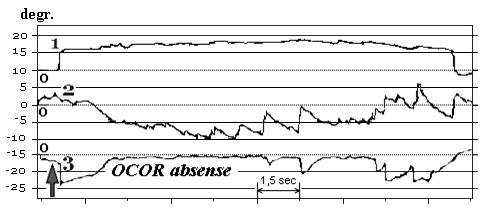
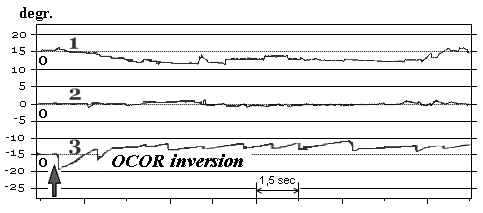
Fig. 11. OCOR without gaze fixation before and after immersion:
1 – horizontal VOG, 2 – vertical VOG, 3 – torsional VOG
↑ – moment of the head tilt to right shoulder
Analysis of OCOR after immersion has shown that 47% of all subjects had significant decrease, absence or inversion of OCOR and positional nystagmus accompanied by an inverted reflex.
Dynamic horizontal vestibular-cervical-ocular reflex (VCOR) and vestibular reactivity (VR)
VCOR was evaluated by gain (gVCOR) as ratio between velocity of the compensatory eyes counter-rolling and velocity of head rotations around the longitudinal axis of the body. VR was evaluated by duration and intensity of the vestibular-invoked nystagmus (time, amplitude and velocity characteristics), which was overlapping (stratifying) the dynamic horizontal counter-rolling of the eyes (during the VCOR test).
Before immersion VCOR was within norm (0.5-0.6) for all subjects. Few subjects had single-strike poorly defined vestibular nystagmus.
After immersion 63% of subjects had no significant changes of gVCOR and 37% of subjects had significantly increased gVCOR accompanied by decreased or atypical OCOR. In this group (with OCOR/VCOR changes) analysis of VR has shown that all nystagmus characteristics except frequency were significantly increased in comparison with baseline up to R+5-6 day after immersion (intensification of the vestibular-induced nystagmus during head rotations around the longitudinal axis of the body). gVCOR and VR’ data are presented in the Table. 1.
Table. 1. Parameters of VCOR and VR before and after immersion
| Parameter | Plane of head movement (rotation) | Day of experiment | |||
| L-7-3 | R+1 | R+3-4 | R+5-6 | ||
| М ± σ | М ± σ | М ± σ | М ± σ | ||
| gVCOR |
Hor. Sag. |
0.77 ± 0.14 0.80 ± 0.11 |
1.10 ± 0.24 * 0.98 ± 0.19 * |
0.90 ± 0.18 0.86 ± 0.20 |
0.79 ± 0.11 0.72 ± 0.15 |
|
VR – TNy, % |
Hor. Sag. |
17.6 ± 6.5 24.1 ± 9.8 |
32.5 ± 10.2 * 39.7 ± 13.4 * |
14.6 ± 9.3 19.0 ± 10.1 |
28.4 ± 8.6* 37.1 ± 10.2 * |
|
VR – FNy, Hz |
Hor. Sag. |
0.43 ± 0.07 0.48 ± 0.09 |
0.50 ± 0.10 0.53 ± 0.14 |
0.44 ± 0.06 0.47 ± 0.10 |
0.48 ± 0.09 0.50 ± 0.10 |
|
VR – ANy, ° |
Hor. Sag. |
5.1 ± 1.4 4.5 ± 2.3 |
9.4 ± 4.1 * 10.8 ± 3.5 * |
4.8 ± 2.0 5.0 ± 2.9 |
11.3 ± 5.9 * 8.7 ± 4.6 * |
|
VR – VNy, °/s |
Hor. Sag. |
15.4 ± 7.2 13.7 ± 4.9 |
25.4 ± 9.2 * 20.7 ± 10.1 * |
22.4 ± 9.1 * 19.7 ± 13.2 * |
14.8 ± 9.0 15.3 ± 7.6 |
* – significant difference with the baseline (L-7-3), p < 0.05
Statistical analysis of all evaluated parameters found positive correlation (0.5…0.9) between parameters of the VT and the MT both during and after immersion. There was positive correlation (0.5…0.8) between vestibular parameters (OCOR, gVCOR/VR) and parameters of the VT on R+1 and negative correlation (-0.6…-0.7) between the same parameters on R+3-4 day after immersion. After immersion there was also found a negative correlation (-0.6…-0.8) between OCOR and gVCOR/VR. However, there was no found correlation between vestibular parameters and parameters of the MT.
Subjective visual vertical (SVV)
Accuracy of perception of the SVV (aSVV) was examined only in 5-day immersion experiment.
Before immersion aSVV in sitting position was within physiological norm (0-2º) for all subjects except two which had aSVV = 3-3.5º. Before immersion the direction of the error of perception for all subjects was leftward – it means that independently of the initial position of visible line (in left or right half-plane on the screen) in the end subjects positioned the line always in left half-plane (Fig. 12).
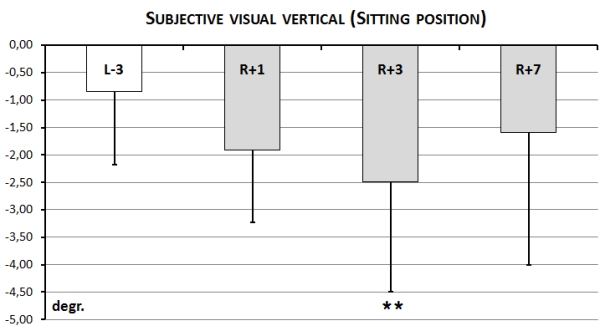
Fig. 12. Accuracy of perception of the subjective visual vertical (aSVV) in a sitting position
After immersion error in perception of SVV in sitting position increased up to 5º and there was a significant different between baseline and aSVV on R+3 day after immersion. It should me mentioned, that for all subjects except one the direction of the error of perception was still leftward..
Before immersion in position lying on the left side (left lateral decubitus) aSVV was within norm = 15-18º. Similar to the sitting position direction of the error of perception was the same (leftward) before, during and after immersion for all subjects except two. During immersion aSVV was lower on all days (I-1, I-3, I-5-6) but there was significant decrease in comparison with baseline only on I-3 day during immersion. On I-5-6 there was a tendency to return of aSVV to the baseline. After immersion aSVV was lower than it was before immersion but significant decrease of aSVV was found only on R+3-4 day. On R+5-6 day aSVV returned to the baseline (Fig. 13.)
It should be mentioned that unlike sitting position decrease of aSVV in left lateral decubitus during and after immersion in comparison with baseline actually means deterioration in the perception of the SVV.
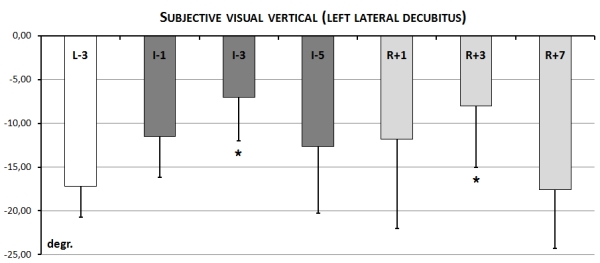
Fig. 13. Accuracy of perception of the subjective visual vertical (aSVV) in left lateral decubitus position
Results presented in this work (statistically analyzed data on 30 participants of the immersion experiment) allow us to state that immersion by changing level and nature of support and proprioceptive afferentation through the central vestibular system (multi-sensory vestibular nuclear complex and its relationship with oculomotor and motor centers) leads to a deterioration in the vestibular, oculomotor and motor responses.
Examinations of the vestibular function which were carried out before and after immersion allowed to register for the first time significant changes in the vestibular system. 47% of subjects participated in the experiment had significant decrease, absence or inversion of OCOR and positional nystagmus accompanied by inverted otolith reflex. 37% of subjects with decreased or atypical OCOR have shown significant increase of gVCOR and increase of VR (intensification of the vestibular-induced nystagmus during head rotations around the longitudinal axis of the body). Accuracy of perception of the SVV was significantly changed on 47% of subjects and there changed correlated with changes of OCOR.
It is believed that the support afferentation in normal conditions has inhibitory effects on the vestibular-oculomotor reflexes. The absence of the support afferentation in immersion, it would seem, should release the vestibular system from this inhibitory control.
However in our studies removal of the support and decrease of the proprioceptive afferentation were accompanied by development of inhibitory effects which, through the efferent nerve fibers from the neurons located in the vestibular nuclear complex, resulted in suppression of the otolith reflex. In turn, suppression of the otolith reflex was accompanied by increase of gVCOR and VR which was proved by significant negative correlation between them. Similar reciprocal relationship between otolith and semicircular canals was also found on cosmonauts after prolonged spaceflights.
Thus, by changing the functioning of the vestibular nuclear complex caused by removal of support afferentation there is a change in nature of central intersensory interactions that can, through efferent connections, also affect the function of the vestibular input.
It is known that mechanism of perception of vertical is based on two components: sensory (intersensory interaction) and motor (skeletal muscle tone). We believe, that support unloading and minimization of the proprioceptive afferentation could lead both to changes in skeletal muscle tone and indirectly (through central mechanisms of the vestibular system) to changes of the vestibular input. Since after immersion there was decrease, absence or inversion of the otolith reflex, it can be assumed that reasons of decrease of accuracy of perception of the SVV both in sitting and lying on side positions were changes in sensory component of perception. The role of the otolith input in accuracy of perception of the SVV is clearly shown in studies when there were examined deaf subjects with no or a drastically reduced otolith reflex. However to understand the leading factor (change in functioning of the vestibular input or change in muscle tone) which affected perception of the SVV we need further research in this direction.
It was found that removal of the support and minimization of the proprioceptive afferentation in immersion has a greater impact on the accuracy of the VT than the accuracy of the motor act of hand (MT) that tracks the movement of the visual stimulus.
It should be noted, that in our studies in normal terrestrial condition in tasks to pursue a predictable movement of visual stimuli with biological feedback the hand motor coordination in VMT was more stable and accurate than in VT without such feedback. Our observations confirm some previous results of other authors in terrestial conditions.
During immersion efficiency ratio (amplitude coefficient) and gain (velocity coefficient) of MT were significantly higher (as well as their coefficients of variation (CV) were significantly lower) in comparison with same parameters of VT. It indicates a higher stability and lower variance of characteristics of the MT in comparison with characteristics of the VT during support-proprioceptive deprivation.
Significant differences were also observed in methods of correction of positional errors of hand and eyes. Eyes had more positional errors which were corrected by corrective saccades. Percent of corrective saccades was significant increased during the whole immersion and until R+3-4 day after.
Hand, unlike the eyes, does not have the mechanism responsible for the production of very fast, accurate and high-speed low-amplitude jerky tracking movements. So in the presence of visual feedback hand is moving toward the target with a little corrective movements. Perhaps this is due to the fact that to generate a hand movement two ways are used: first – direct as effector commands and proprioceptive feedback signals, and second – indirect using visual information.
It should me mentioned, that latency of the MT in task of fixation and holding a “hand” target on the jerky (saccadically) moving stimulus was more than 1.5-2 times higher than latency of the VT in the same task. That is, the eyes reach their target sooner than the hand, which was confirmed by other researchers. It indicates a longer reflective pathway (in comparison with eyes saccades) of hand effector command to move a hand from one side position to another, i.e. “stimulus – eyes” reaction time (loopback) is lower (shorter) than “stimulus – hand” reaction time (loopback).
Total reaction time of VT was significantly higher than one of MT during the whole immersion and on R+1 day after. From our point of view, significant increase of total reaction time of VT in comparison with MT was caused by process (strategy) of saccadic approximation which replaced smooth tracking.
Immersion allowed us not only to determine the natural changes in the characteristics of VMT, but also to identify individual differences in these characteristics, which indicates a different role of the support-proprioceptive factor in mechanisms of intersensory interactions and in particular in visually-mediated interaction between hand and eyes motor system.
The nature of changes of VMT also related on duration of sensory deprivation: the 5-6 day immersion was not sufficient to complete a period of adaptation to the support unloading.
Statistical analysis of results obtained in immersion experiment allowed us to find a positive correlation (0.5…0.9) between characteristics of the VT and MT both during and after immersion. There was found a positive correlation (0.5…0.8) between vestibular parameters (OCOR, gVCOR/VR) and parameters of the VT on R+1 and negative correlation (-0.6…-0.7) between the same parameters on R+3-4 day after immersion. After immersion there was also found a negative correlation (-0.6…-0.8) between OCOR and gVCOR/VR. However, there was no found correlation between vestibular parameters and parameters of the MT.
Conclusion
The results on the effects of immersion on the state of vestibular reflexes and “eyes-hand-stimulus” motor coordination allowed us to clarify the understanding of organization of multisensory provision of spatial orientation during support unloading and minimization of proprioceptive afferentation.
Examinations of the vestibular function (VF) which were carried out before and after immersion allowed to register for the first time significant changes in the vestibular system. 47% of subjects participated in the experiment had significant decrease, absence or inversion of OCOR and positional nystagmus accompanied by inverted otolith reflex. 37% of subjects with decreased or atypical OCOR have shown significant increase of gVCOR and increase of VR (intensification of the vestibular-induced nystagmus during head rotations around the longitudinal axis of the body). Accuracy of perception of the SVV was significantly changed on 47% of subjects and there changed correlated with changes of OCOR.
Along with found correlation between parameters of the VT and MT, between VF and VT there was no found correlation between VF and MT. It was shown, that support unloading and minimization of the proprioceptive afferentation has a more pronounced effect on the accuracy of the VT than the accuracy of the MT. MT was more accurate and stable than VT for all subjects.
Comparative analysis of results obtained after spaceflight and after immersion shows that similarity of changes of VF and VMT. However after immersion these changes are less pronounced and the process of recovery to the baseline takes much less time (5-6 days for all immersion subjects). Duration of the postflight readaptation is 10 days for 64% of examined cosmonauts and 14-18 days for the remaining 36% of cosmonauts.